
6.4.3. Сокращение числа потенциальных работников
В России к трудоспособному возрасту принято относить мужчин от 16 до 60 и женщин от 16 до 55 лет. В международной практике к этой категории относят обычно и мужчин, и женщин в возрасте от 15 до 65 лет. В настоящем докладе приводятся показатели как по первому («российский критерий»), так и по второму («международный критерий») варианту.
Численность населения в трудоспособном возрасте (далее для краткости — трудоспособного населения) сокращается по всем вариантам прогноза (табл. 6.8).
Таблица 6.8. Медианное значение численности трудоспособного населения по разным вариантам прогноза, Россия, 2000-2050 гг., млн. человек
|
Год |
Критерии выделения трудоспособных |
|||
|
Российский |
Международный |
|||
|
Инерционный вариант |
Вариант с неизменной численностью населения |
Инерционный вариант |
Вариант с неизменной численностью населения |
|
|
86,5 |
86,5 |
100,8 |
100,8 |
|
|
86,1 |
89,8 |
104,2 |
||
|
73,8 |
82,4 |
99,9 |
||
|
79,8 |
||||
|
58,1 |
79,2 |
72,1 |
96,8 |
|
|
47,7 |
74,4 |
61,1 |
92,5 |
|
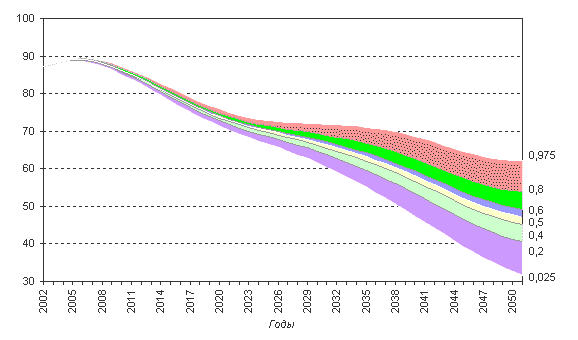
Рисунок 6.8. Численность населения в трудоспособных возрастах (российский критерий) при разных доверительных интервалах, в млн. человек. Инерционные сценарии прогноза
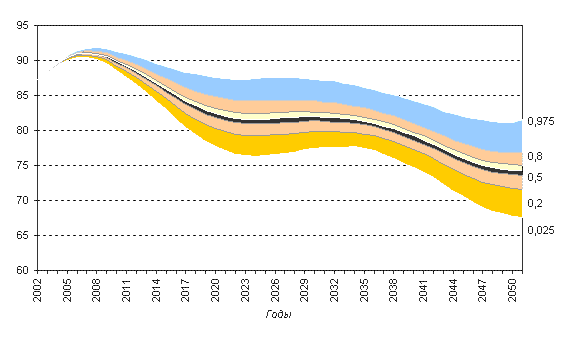
Рисунок 6.9. Численность населения в трудоспособных возрастах (российский критерий) при разных доверительных интервалах, в млн. человек. Сценарии с постоянной численностью населения
Численность трудоспособного населения сокращается по всем вариантам прогноза. Однако, разумеется, при сохранении постоянной численности населения, его трудоспособная часть, сокращаясь в результате старения, убывает не так быстро, в период примерно между 2020 и 2035 годами почти не меняется и лишь затем переходит в фазу нового падения. При этом при использовании международного критерия выделения трудоспособного населения это падение оказывается менее крутым — всего на 8% по сравнению с 2000 годом против 14% при использовании российского критерия.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изучение соотношения в подходах рыб ранней и поздней темпоральных форм предпринято с 1990 г. (рис. 2), тем самым охватывая последний период высокого уровня запасов горбуши в регионе, начавшийся с конца 1980-х гг. и завершившийся к середине 2010-х гг. (Kaev, Irvine, 2016). В этот же период (с 1990 г.) были начаты исследования горбуши на Кунашире. Быстрый темп прироста уловов на этом острове в чётные годы за счёт увеличения численности рыб обеих темпоральных форм привёл к доминированию с 1994 г. этой генеративной линии по уровню запаса (рис. 2а). В годы наибольшей численности горбуши (1998 и 2000) доля ранней формы в подходах была выше. В 2002 г. наметилась тенденция снижения доли ранней формы вследствие уменьшения её численности, в то время как численность поздней формы даже несколько возросла, после чего последовало резкое сокращение численности рыб уже обеих темпоральных форм. В последующие годы доминирование по доле рыб в подходах в чётные годы перешло к поздней форме на фоне общего низкого уровня запаса. По линии нечётных лет начиная с 1993 г. наблюдалось устойчивое доминирование по доле в подходах рыб ранней формы при сравнительно низком в среднем уровне запаса. Его заметный рост последовал с 2001 г. в основном за счёт увеличения численности ранней формы. После достижения пика в 2007 г. произошло резкое снижение запасов горбуши, особенно её ранней формы, в результате чего на её долю в 2011 г. пришлось менее половины возврата. В 2013–2017 гг. соотношение форм не установлено ввиду прекращения биологических сборов и отсутствия данных по динамике подходов горбуши из-за ведённых ограничений на её промысел при критически малой численности рыб в возвратах.
Рис. 2.
Динамика численности ранней ( ) и поздней (
) и поздней ( ) темпоральных форм горбуши Oncorhynchus gorbuscha по линиям нечётных и чётных лет в 1990–2017 гг.: а − Кунашир, б – Итуруп, в − юго-восточное побережье Сахалина; (∙∙∙∙∙∙) – суммарная численность обеих форм, (–––) – доля ранней формы.
) темпоральных форм горбуши Oncorhynchus gorbuscha по линиям нечётных и чётных лет в 1990–2017 гг.: а − Кунашир, б – Итуруп, в − юго-восточное побережье Сахалина; (∙∙∙∙∙∙) – суммарная численность обеих форм, (–––) – доля ранней формы.
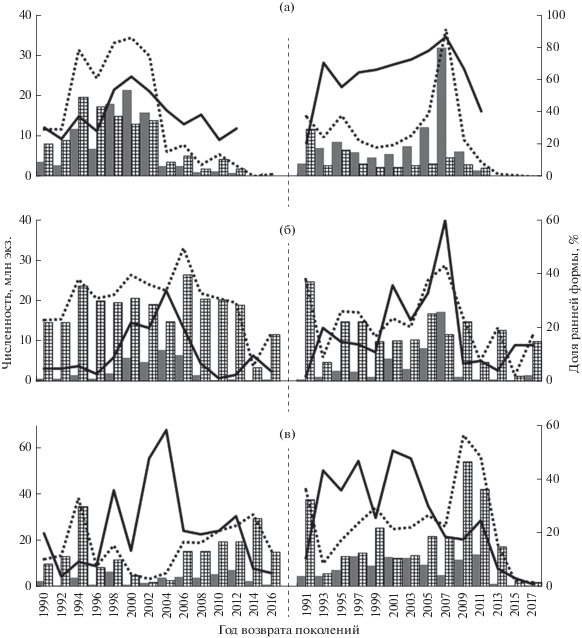
На Итурупе доминирование по уровню запаса также с 1994 г. перешло к генеративной линии чётных лет. По этой линии с 1998 г. наблюдалось увеличение в подходах рыб ранней формы, в то же время этот прирост незначительно повлиял на изменение общей численности горбуши вследствие слабого присутствия в подходах рыб этой темпоральной формы – от 9 до 34%. Достижение исторического максимума уровня запасов в 2006 г. было обусловлено в основном за счёт роста численности поздней формы, в то время как численность ранней формы в этом году обозначила тенденцию к снижению, а в 2008 г. резко сократилась (рис. 2б). Высокий уровень запасов стада продолжал сохраняться до 2012 г. за счёт стабильно больших возвратов рыб поздней формы, которые затем также пошли на спад. Кратковременное увеличение доли рыб ранней формы вследствие резкого снижения численности поздней формы в 2014 г. не совсем вписывается в тенденцию, оно было связано с массовым выловом курильской горбуши (в основном её поздней формы) в водах южной части Сахалина, видимо, из-за миграционного «сбоя” (Каев, Животовский, 2016). По линии нечётных лет рост численности ранней формы в первом десятилетии XXI в. привёл к максимуму уровня запасов горбуши этой генеративной линии в 2007 г., после которого произошло резкое сокращение численности рыб этой темпоральной формы и снижение общего уровня запасов, в том числе и за счёт уменьшения численности рыб поздней формы.
На юго-восточном побережье Сахалина изменение запасов горбуши в чётные годы определялось в основном возвратами рыб поздней формы, так как доля ранней формы составляла от 4 до 36%; лишь в 2002 и 2004 гг. она возросла до 48 и 59%, когда численность обеих форм была на минимальном уровне (рис. 2в). Тем не менее определённое значение в формировании двух «волн” состояния запасов горбуши в рассматриваемые чётные годы принадлежало и ранней форме, численность которой резко сокращалась в преддверии общего снижения запасов. По генеративной линии поколений нечётных лет ранняя форма горбуши, на долю которой в 1991–2011 гг. приходилось в среднем 32%, имела большее значение в формировании динамики запасов рыб этого вида. Так, при их историческом максимуме в 2009 и 2011 гг. численность ранней формы также была на уровне максимальных значений, а при последующем резком сокращении запасов численность этой формы в первую очередь тоже снизилась до минимума.
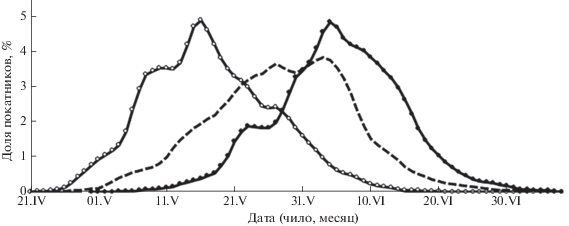
Рассмотрим, как сказывалось на соотношении темпоральных форм в возвратах горбуши действие экстремальных факторов среды. Прежде всего, для проведения такого анализа следует уточнить, что по срокам нереста горбуши в рассматриваемых районах существенных расхождений нет. Производители ранней формы в реках южных Курильских о-вов нерестятся в основном в августе и в первой половине сентября, а поздней формы – в сентябре и в первой половине октября; на юго-восточном побережье эти сроки смещаются на более ранние даты ориентировочно на 5 сут. Иначе обстоит ситуация с покатной миграцией, динамика которой схожа с динамикой нерестового хода горбуши, определяемой соотношением рыб ранней и поздней темпоральных форм (Каев, 2012; Каев, Ромасенко, 2013). В то же время отсутствует корреляция между датами середины нерестового хода и последующей покатной миграцией, что связано, видимо, с различиями в скорости развития эмбрионов при разных температурных режимах (Каев, Ромасенко, 2002). Действительно, в рассматриваемых районах наиболее поздние сроки миграции основной массы молоди характерны для юго-восточного побережья Сахалина, где в зимний период велико влияние материковых холодных масс воздуха, а наиболее ранние – для Кунашира, на котором свойственные для южнокурильских островов мягкие зимы сочетаются с влиянием тёплого течения Соя, омывающего этот остров (Каев, Ромасенко, 2017). Кроме того, более ранние сроки ската основной массы молоди на Кунашире обусловлены наибольшим представительством ранней темпоральной формы. В соответствии с этим в конфигурации кривой, характеризующей динамику покатной миграции молоди из рек Кунашира, хорошо просматривается положительная асимметричность, тогда как кривые Итурупа и юго-восточного побережья Сахалина имеют отрицательную асимметричность (рис. 3).
Рис. 3.
Среднемноголетняя динамика покатной миграции молоди горбуши Oncorhynchus gorbuscha в разных районах воспроизводства: (—⚪—) – Кунашир, (—) – Итуруп, (—⚫—) – юго-восточное побережье Сахалина.
На Кунашире (рис. 4а) обильный возврат горбуши в 2007 г., на первый взгляд, не соответствует штормовым ветрам при нагуле молоди этого поколения. Однако первый шторм (2 мая) произошёл до начала массового ската молоди из рек, а второй (1 июня) воздействовал в основном на поздно скатившуюся молодь, так как подросшие ранее скатившиеся мальки уже не входят в группу риска по отношению к действию данного фактора (Каев, 1992). Существенное снижение численности следующего поколения по генеративной линии нечётных лет было обусловлено паводком (25 сентября), когда отнерестились производители ранней формы и бóльшая часть производителей поздней формы, а при нагуле молоди выжившей части этого поколения штормовые условия (21 мая) воздействовали в основном на первую половину мигрантов. То есть в обоих случаях давлению среды подверглась в основном ранняя форма горбуши, что и обусловило резкое снижение численности рыб этой формы в 2009 г. Воздействие штормовых ветров на нагул молоди следующего поколения способствовало усилению депрессии этой генеративной линии горбуши, особенно её ранней формы. В рассматриваемый период поколения генеративной линии чётных лет уже находились в состоянии депрессии, которая продолжала сохраняться под действием данных факторов, воздействовавших в большей мере опять-таки на воспроизводство ранней формы горбуши. Череда циклонов в последующие годы углубила депрессивное состояние запасов этого стада по обеим генеративным линиям, так как в дополнение к их действию появился ещё один мощный отрицательный, уже внутренний, фактор: величина возвратов горбуши стала недостаточной для заполнения нерестилищ. Есть надежда на увеличение численности последующих поколений горбуши в основном за счёт возврата рыб поздней темпоральной формы, так как на результатах нереста рыб-родителей ранней формы продолжали отрицательно сказываться паводки.
Рис. 4.
Численность возврата ранней и поздней темпоральных форм горбуши Oncorhynchus gorbuscha по линиям нечётных и чётных лет и случаи воздействия экстремальных факторов среды в период нереста их родителей и нагула молоди в прибрежных морских водах: а − Кунашир, б – Итуруп, в − юго-восточное побережье Сахалина; (◇, ◆) – паводковые осадки, повлиявшие на результаты нереста производителей соответственно ранней формы и обеих форм, (△,  , ▲) – штормовые ветры в период нагула молоди на прибрежном морском мелководье при скате соответственно первой, второй и последней трети молоди из рек; обозначения форм см. на рис. 2.
, ▲) – штормовые ветры в период нагула молоди на прибрежном морском мелководье при скате соответственно первой, второй и последней трети молоди из рек; обозначения форм см. на рис. 2.
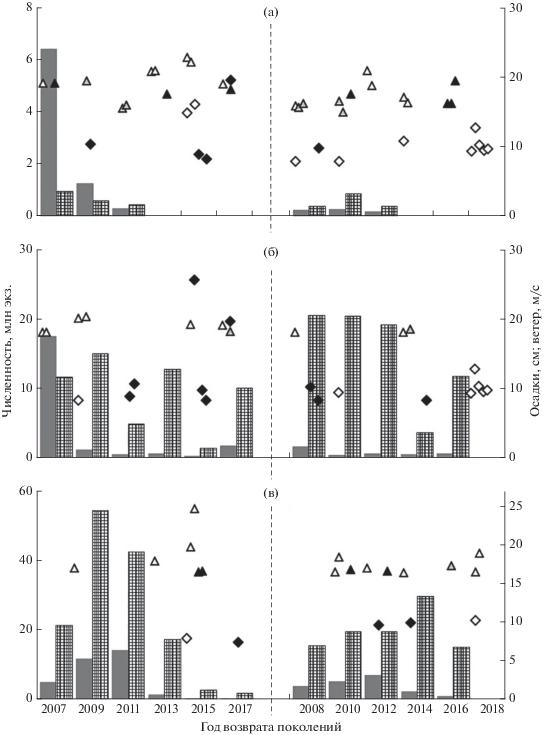
На Итурупе (рис. 4б) для поколения горбуши возврата 2007 г. сложились, видимо, благоприятные условия воспроизводства ранней формы, на которую не повлияли штормовые ветры (11 и 13 мая), наблюдавшиеся до начала массового ската молоди из рек. Однако такие ветры и частично паводок в начале нереста в полной мере отразились на воспроизводстве ранней формы следующего поколения, её численность резко сократилась по отношению к таковой родительского поколения. Слабый возврат рыб обеих темпоральных форм в 2011 г. был обусловлен воздействием двух паводков, один из которых наблюдался в конце нереста, а второй – после его завершения. Некоторое снижение численности горбуши в 2013 г. по отношению к таковой в 2009 г. связано в основном с частичным дефицитом в 2011 г. производителей на нерестилищах (1235 тыс. против 1423 тыс. экз. в среднем за предыдущие цикличные годы), хотя в отношении ранней формы можно полагать и ухудшение условий нагула её молоди. Совместное действие экстремальных факторов среды привело к ещё большему сокращению численности обеих темпоральных форм в 2015 г., в том числе и к слабому заходу производителей на нерестилища (637 тыс. экз.). Тем не менее в результате благоприятных условий нагула рыб в морских водах (индекс выживания 6.35 против 3.58% четырёх предыдущих цикличных поколений) в 2017 г. отмечен прирост численности горбуши, в большей мере её поздней формы, так как действию экстремальных факторов среды частично подверглось воспроизводство только ранней формы (это воздействие на Итурупе могло быть меньшей силы, чем оно оценено по данным МП Южно-Курильск).
Численность поколений горбуши генеративной линии чётных лет, особенно ранней темпоральной формы, на фоне действия экстремальных факторов среды на Итурупе рассматривается для периода уже произошедшего её снижения с 2006 г. Численность рыб поздней формы незначительно изменялась в 2008, 2010 и 2012 гг., а численность ранней формы продолжила своё снижение в 2010 г. вследствие паводка при нересте родителей, а в 2012 г., видимо, из-за возникшего дефицита производителей этой темпоральной формы на нерестилищах вследствие её минимального представительства в подходах в 2010 г. В 2014 г. можно было ожидать дальнейшего сокращения численности обеих темпоральных форм, но не такого значимого, как это неожиданно произошло у поздней формы, так как штормовые ветры действовали в основном на первую половину скатившейся из рек молоди. Причиной столь непропорционального снижения численности рыб поздней формы стало перемещение части рыб к южному побережью Сахалина, о чём упоминалось выше. Сравнительно небольшую численность рыб поздней формы в 2016 г. и практически отсутствие её прироста у ранней формы в сравнении с предыдущим цикличным поколением только частично можно объяснить дефицитом производителей на нерестилищах. Вероятно, в большей мере здесь сказалось постепенное ухудшение условий нагула рыб в морских водах в ряду поколений с возвратом рыб в 2010, 2012 и 2016 гг., судя по значениям индекса их выживания (соответственно 6.39, 6.10 и 5.92%). Рекордно большой скат молоди из рек в 2017 г. (Каев и др., 2017) даёт надежду на восстановление высокой численности горбуши по генеративной линии чётных лет, в том числе и её ранней формы, так как не отмечено значимого влияния на динамику ската молоди серии предполагаемых паводков с 17 августа по 9 сентября при нересте родителей (по данным МП Южно-Курильск).
На юго-восточном побережье Сахалина (рис. 4в) рекордные возвраты горбуши в 2009 и 2011 гг. были обеспечены быстрым приростом численности рыб обеих темпоральных форм. Штормовые условия при нагуле молоди на прибрежном морском мелководье одного из этих поколений (1 мая) не помешали развитию этого процесса, так как наблюдались до начала массового ската молоди из рек. Существенное снижение численности в 2013 г. следует рассматривать как результат развития каких-то иных природных процессов, которые сильнее отразились на формировании численности ранней формы, ибо штормовые ветры (13 мая) при нагуле ещё только небольшой части скатившихся мальков не могли обусловить такое снижение численности ранней формы. А вот катастрофическое снижение численности горбуши в следующем цикличном году, вплоть до почти полного исчезновения в подходах рыб ранней формы, произошло, видимо, по причине частых штормовых условий на прибрежном морском мелководье, воздействовавших в основном на нагул первых двух третей скатившихся из рек мальков (16−17 мая и 11−12 июня). Депрессия запасов горбуши обеих темпоральных форм ещё более углубилась в 2017 г. вследствие возникшего дефицита производителей при нересте родительского поколения (983 тыс. против 2346 тыс. экз. в среднем для пяти предыдущих цикличных поколений) и воздействия паводка на результаты их нереста после его завершения. Генеративная линия чётных лет является рецессивной по уровню запаса, поэтому действие экстремальных факторов среды следует рассматривать в её отношении лишь как усугубляющее и так менее благоприятные условия воспроизводства. Можно полагать, что эти факторы сдержали наметившийся рост численности обеих темпоральных форм в 2008, 2010 и 2012 гг. О приросте численности рыб поздней формы в 2014 г. как следствия перераспределения горбуши упоминалось выше. Судя по результатам учёта покатной молоди (Каев и др., 2017), возможно увеличение численности горбуши за счёт возврата в этом поколении рыб поздней формы, в то время как численность рыб ранней формы останется низкой вследствие негативного влияния штормов на нагул молоди. Из вышеизложенного следует, что между разными темпоральными формами горбуши существуют различия по динамике численности. Годы с высокой или низкой численностью рыб совпадают по группам лет, однако периоды подъёма численности ранней формы в целом короче, что особенно хорошо заметно при резком падении уровня запаса горбуши, происходившего в последние годы. Снижение численности ранней формы наступало либо раньше на одно–два поколения, либо было более интенсивным при синхронном (в смежные годы) развитии этого процесса, что хорошо иллюстрирует уменьшение доли рыб ранней формы в подходах. Зачастую снижение численности каждой из темпоральных форм было обусловлено действием экстремальных факторов среды, но в некоторые годы более интенсивное снижение численности ранней формы определялось какими-то иными причинами. Такие изменения численности ранней формы вполне соответствуют заключению о её более высоком и изменчивом уровне смертности, с чем согласуется бо́льшая относительная плодовитость самок этой формы (Каев, 2012). Рассмотренные различия в динамике численности темпоральных форм могут быть связаны как с нерестом производителей в разное время и на разных нерестилищах, так и с условиями нагула молоди в морском прибрежье при разных сроках ската из рек.
Морфобиологические и экологические различия вкупе с особенностями динамики численности указывают на определённую обособленность темпоральных форм горбуши, которые сравнительно широко распространены по ареалу размножения (Иванков, 2011). В то же время следует осторожно относиться к заключению, что у горбуши эти темпоральные формы имеют ранг экологических подвидов (Иванков, Иванкова, 2017), учитывая высокую вероятность скрещивания между ними при существующем широком перекрытии их сроков миграции и мест нереста. Критически следует воспринимать также предложенную общую для горбуши и кеты O. keta схему внутривидовой эколого-темпоральной дифференциации, в соответствии с которой эти виды подразделяются на эколого-темпоральные расы – русловую и ключевую, а последняя включает экотипы мелких рек, крупных рек и озёрный (Иванков, Иванкова, 2017). Поздняя форма горбуши весьма условно вписывается в ключевую расу, так как лишь только отдельные её производители встречаются на ключевых чашах в бассейнах рек. В целом же нерестилища ранней и поздней формы горбуши «привязаны» к подрусловому потоку, различия между ними заключаются только в их разной в среднем удалённости от устья. Возможно, по этой причине горбушу первоначально точно по такой же схеме объединяли с симой O. masou, а кету – с неркой O. nerka (Иванков, Иванкова, 2013). Непонятным остаётся выделение в этой схеме у горбуши и кеты озёрного экотипа, включающего по две темпоральные популяции (Иванков, Иванкова, 2017). При миграции кеты в озёрно-речные системы южных Курильских о-вов, где описан озёрный экотип данного вида (Иванков, 1984, 1985; Каев, Ромасенко, 2010), действительно наблюдаются две группы рыб, различающиеся по высоте тела и степени выраженности внешних брачных изменений. Но это не темпоральные популяции, а рыбы разных экотипов – озёрного (нерестящегося в местах выхода грунтовых вод в ложе озера) и речного (нерестящегося на кетовых ключевых нерестилищах в небольших реках, впадающих в озеро). Последние по генетическим маркерам сходны с рыбами даже из сравнительно удалённых рек острова, а не с рыбами озёрного экотипа в данной озёрно-речной системе (Каев и др., 2008). В эти же системы заходит горбуша, мигрируя далее в реки. К настоящему времени нет свидетельств о её нересте непосредственно в озёрах, которые являются лишь зоной транзита.
Таким образом, в настоящее время можно говорить только о существовании темпоральных форм горбуши, в определённой степени различающихся по срокам и местам нереста на русловых нерестилищах в одних и тех же реках, биологическим показателями рыб (различия по соотношению длин самцов и самок, относительной плодовитости самок, степени полового созревания при заходе в реки) и срокам покатной миграции (Каев, Ромасенко, 2017), что и создаёт предпосылки для особенностей динамики численности каждой из форм. Как правило, снижение численности ранней формы начинается раньше или происходит интенсивнее, при этом причины снижения не всегда удаётся установить при ограниченном наборе изучаемых параметров среды в ходе мониторинга. Важно, что ранняя форма быстрее реагирует на изменения условий воспроизводства, в то время как в силу её малочисленности общая численность горбуши может оставаться при возврате последующих одного−двух поколений на прежнем уровне. Поэтому появление признаков депрессивного состояния у ранней формы может служить индикатором предстоящей общей депрессии запасов горбуши в данном районе.



